Accession Data:
Brugmansia pittieri (Saff.) Moldenke
- Common Name: 'Culebra'
- Family: Solanaceae Juss.
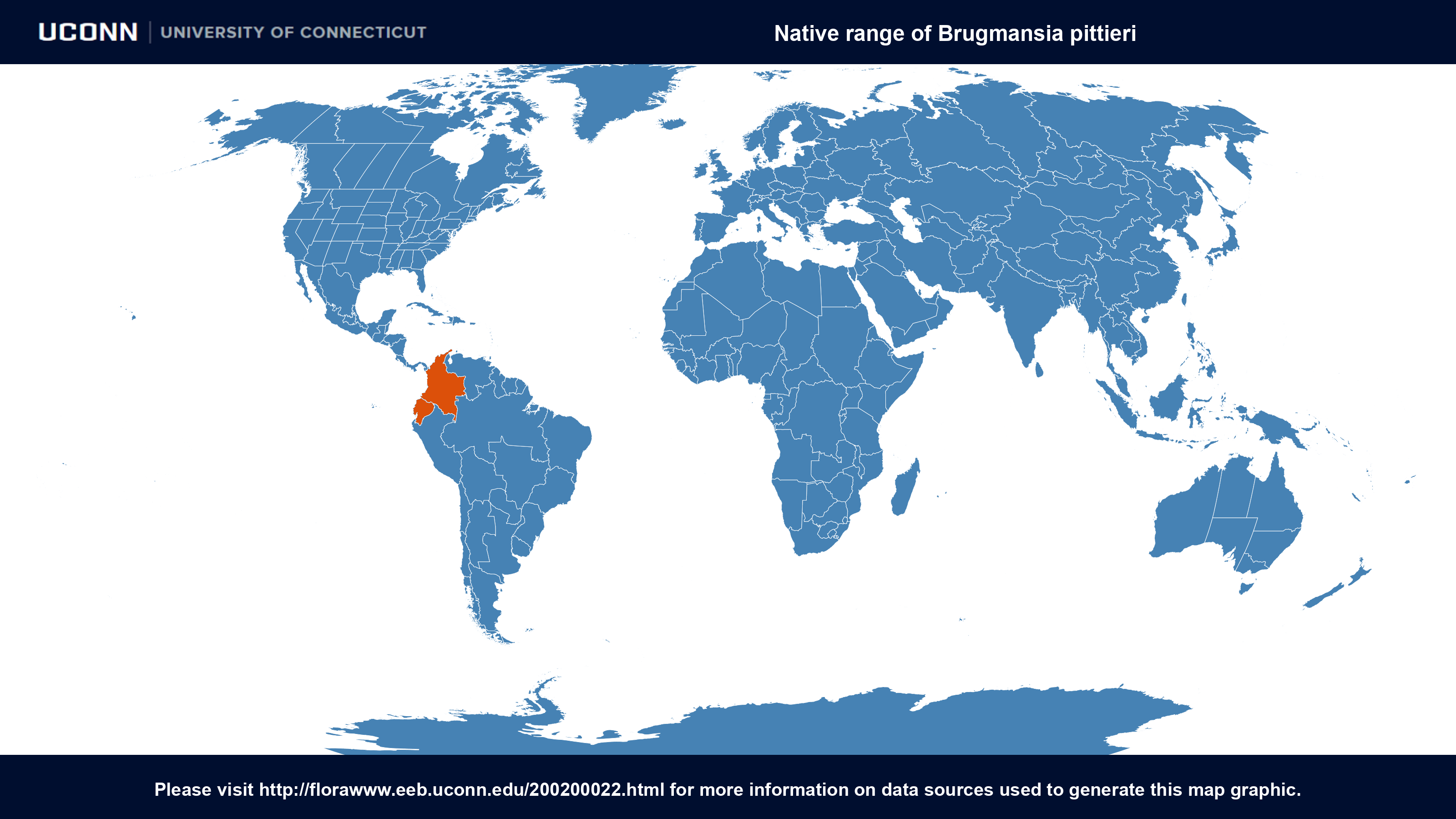
- Country of Origin: Colombia - high and geographically isolated Indian village of Sibundoy
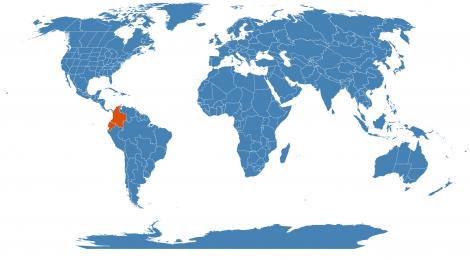
- Habitat: forested slopes of the mountains to the south of the valley of Sibundoy
- Description: Originally classified as Methysticodendron amesianum by R.E. Schultes in 1955 (Bot. Mus. Leaf., 17, 1, 1955)
Description of Methysticodendron Amesianum- R. E. Schultes in Bot. Mus Leafl. Harvard Univ. 17 (1955) 2.
Tree up to 25 feet tall, with short, densely leafy branches. Leaves membranaceous, dark green, narrowly linear-ligulate, apically acuminate, basally gradually attenuate (very often without distinct petiole), marginally commonly subundulate or undulate, in adult state 20-26 cm long, 1.3-2.0 cm wide; central nerve strong, beneath strongly elevated, secondary nerves almost parallel, somewhat conspicuous, towards margin arcuateanastomosing; both surfaces minutely and irregularly (near veins denser) pilose.
Flowers up to 28 (mostly more or less 23) cm long apically 10-13 cm in diameter in bud, at dusk and night very strongly and sweetly fragant. Calyx spathaceous, green, papyraceous or membranaceous, 2- to 5-parted, with acute teeth, tubiform for half its length, 3/5 as long as corolla (up to 13 cm long), both surfaces very minutely and irregularly pilose. Corolla divided for 3/5-4/5 its length, with lobes usually 5 (but abnormally 4 or 6), membranaceous, white, spreading, spathulate or almost spathulately rhombiform, apically long acuminate, circinnate, entire, upper half strongly inflexed, 14-16 cm long, towards apex of tube usually 6mm. (but in widest part 2.5-3.3 cm) wide, near base very remotely and irregularly pilose within and without, with 3 conspicuous, strongly parallel veins; tube rather densely white-pilose within.
Stamens usually 5, shorter than corolla lobes; with terete filaments, more or less 5 cm long, 2 mm in diameter at base, apically strongly twisted and lightly bent, pilose except at apex; anthers yellow, 2.7-3 cm long, 3-4 mm in diameter. Ovary sub-cylindric, conspicuously 3-sulcate, glabrous, up to 1.5 cm long, 0.5 cm in diameter, gradually produced into styles. Styles glabrous, up to more or less 9 cm long, 2.5 mm in diameter, apically slightly clavellate and strongly flattened, with stigmatic tissue on very apex and along adges up to 1-1.5 cm in decurrent form, black (when dry) and papillose. Appendix (rudimentary style?) like styles arising from central part of ovary, up to 5.5 cm long, accompanied by 1 or 2 subclavellate, fleshy appendices 2-5 mm long. There are no herbarium collections of any species of this genus made from confirmed wild plants. No botanist specialising in this genus has ever reported seeing wild plants of any species. (Verbal) Reports by non-specialist botanists of the occurrence of ‘wild’ plants are either misidentifications (usually of Datura), or misinterpretation of remnants or localised escapes from cultivation, usually along creeks and occurring by vegetative propagation from stem fragments. In all such instances investigated in Ecuador and Colombia, the plants are of the anthropogenic hybrid Brugmansia x candida (Hay et al. 2012: 172-177). It is quite clear that such instances do not represent self-sustaining sexually reproducing populations.4
The complete lack of evidence of fruit dispersal or spontaneous seedlings, combined with the presence of large numbers of fruits containing viable seed, suggests their dispersers are extinct. Hence, all the species should best be regarded as extinct in the wild.4
- Uses: This plant is known only from the valley of Sibundoy where it is employed by the Inga and Kamsá Indians in their witchcraft. Its use in each tribe is restricted to several witch-doctors, and these practitioners grow the plant with special care in the neighbourhood of their huts. The wild habitat of the species is said to be the forested slopes of the mountains to the south of the valley of Sibundoy, especially the slopes of the botanically almost completely unknown Cerro Patascoy. When in cultivation, the plant is apparently the hereditary property of certain families, as the sorcerers pass it on to the eldest son together with the secrets attending its use.
- IMPORTANT NOTE: Plant Uses are for informational purposes only. EEB Greenhouses assume no responsibility for adverse effects from the use of any plants referred to on this site. Always seek advice from a professional before using any plant medicinally.
- USDA Zone: 8a-11
Accession Data:
- Accession # 200200022
- Source: Rich Sanders
- Provenance: Cuttings from Richard Sanders Feb2002. Original material from Andre Hodnik, France ex Sibundry Valley, SW Columbia (only known wild source of this clone)
- Accession Date: 02-11-2002
- Bench: 2116 - MED:Mattoral B
- Currently: active - healthy
- Qty: 2 confirmed on 03-27-2025
- Restrictions:
- Poisonous Plant Parts - Not for Human Consumption
tropane alkaloids - fatally toxic. If ingested, immediately call the Poison Control Center. - IUCN Red List: Endangered Species - Extinct in the Wild
- BGCI Limited Ex Situ Distribution - Plants do not leave building without Manager approval!
- Poisonous Plant Parts - Not for Human Consumption
Classification:
- Division: Magnoliophyta
- Class: Magnoliopsida
- SubClass: euasterid I
- Order: Solanales
- SubOrder:
- Family: Solanaceae
- SubFamily: Solanoideae
- Tribe: Datureae
- SubTribe:
Flowering Data:
This accession has been observed in bloom on:| Year | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2025 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2024 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2023 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2022 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2021 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2020 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2019 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2018 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2017 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2016 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2015 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2014 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2013 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2012 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2011 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2010 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2009 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2008 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2007 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2006 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2005 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2004 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
References (internal):
References (external):
- Personal communication with Rich Sanders, who provided the cuttings for us.
- Methysticodendron amesianum - posted at Lycaeum Website - accessed 26 April 2002.
- The Plant List (2013). Version 1.1. Accessed 24 June 2015.
- Hay, A. 2014. Brugmansia aurea. at IUCN Red List of Threatened Species. Version 2015.2. Last accessed on Thursday, December 15, 2016.
- Poisonous Plants at University of California. Last accessed on Wednesday, July 05, 2017.
data regenerated on Wed, 14 May 2025 10:30:11 -0400 [bcm v4.0]
Images:

Additional images for this accession:
Click on thumbnails to enlargeCurrent Accessions in the Solanaceae
Subfamily Browallioideae
Tribe Cestreae
Subfamily Nicotianoideae
Subfamily Petunioideae
Subfamily Solanoideae
Subfamily Solanoideae
Tribe Capsiceae
Subfamily Solanoideae
Tribe Datureae
Subfamily Solanoideae
Tribe Hyoscyameae
Subfamily Solanoideae
Tribe Juanulloeae
Subfamily Solanoideae
Tribe Lycieae
Subfamily Solanoideae
Tribe Physaleae
- Iochrominae: Iochroma cyaneum Royal Blue

- Iochrominae: Iochroma fuchsioides
- Withaninae: Withania riebeckii W/C
- Withaninae: Withania somnifera
Subfamily Solanoideae
Tribe Solaneae
- Jaltomata quipuscoae
- Solanum conocarpum
- Solanum conocarpum
- Solanum ensifolium
- Solanum ensifolium W/C
- Solanum seaforthianum
- Solanum wendlandii
 = indicates flowering in past 14 days
= images available for this accession
= map available for this accession
= indicates flowering in past 14 days
= images available for this accession
= map available for this accession
 = accession added within past 90 days
= accession added within past 90 days